ПРОСТРАНСТВЕННО-СОЦИАЛЬНАЯ ОРГАНИЗАЦИЯ: ГОРОДСКИЕ БЕЗНАДЗОРНЫЕ СОБАКИ ИЗУЧЕНЫ ВНОВЬ Энрике Фонт (Enrique Font , факультет биологических наук, Университет Валенсии, Испания).
Spacing and Social Organization: Urban Stray Dogs Revisited.
(Журнал Applied Animal Behaviour Science, 17 (1987), p. 319-328.)
( Эта работа, прежде всего, примечательна тем, что в ней обсуждается вопрос о социальности безнадзорных (в том числе и бездомных) собак в городских условиях. Как известно, ранние (фактически самые первые в мировой научной практике, датируемые 1970-80-ми годами) работы американских исследователей по количественной экологии безнадзорных собак в городских условиях показали их очевидную «асоциальность» - то есть преимущественно одиночный образ жизни, отсутствие постоянных семейных групп – стай. Эти выводы, полученные в конкретных условиях крупных американских городов, оспариваются в настоящей работе европейского автора, наблюдавшего стаи в испанской Валенсии. Автор делает вывод, что социальность собак зависит от конкретных условий обитания. Вместе с тем, он подвергает критике метод, использованный американцами для выявления стай, справедливо указывая на то, что долгосрочные этологические наблюдения и оценка многих поведенческих параметров предпочтительнее использования формальных математических моделей, в которых в расчет берется лишь расстояние между особями. Однако, следует заметить, что большинство безнадзорных собак в американских городах действительно могло быть одиночными, так как там значительную долю составляли владельческие собаки на бесконтрольном «самовыгуле». Такие собаки не склонны формировать постоянные стаи, так как в значительней мере социализированы на своих владельцев, к дому которых они постоянно возвращаются, где получают пищу и т.д. Субпопуляция же бездомных собак находилась под постоянным прессом службы отлова (Animal Control), не позволявшим сформировывать стаи (в том числе и в результате самовоспроизводства, когда подросшие щенки остаются с самкой). Преобладающая часть бездомных собак была бывшими владельческими. Конечно, в других условиях – например, при менее интенсивном контроле, наличии пустырей, огороженных нежилых территорий - собаки получают возможность сформировывать стаи и в городских условиях, хотя прямых преимуществ это, видимо, не дает. – В.Р.)
Были изучены различные аспекты это-экологии популяции бродячих (безнадзорных) собак в Валенсии. Популяционные плотности между 127 до 1304 безнадзорных собак на кв. км соответствуют интервалам плотностей, полученных для других популяций (имеются в виду американские работы, такие огромные плотности, видимо, результат учета всех собак – прежде всего владельческих безнадзорных; в настоящей работе при общем учете также не делалось различия между бездомными и безнадзорными владельческими собаками – В.Р.) . Соотношение самцы-самки: 2 к 1 – также соответствует более ранним работам. Наблюдения поведения показали, что эти собаки иногда формируют группы с иерархией доминирования и совместной защитой территории. Исходя из стабильности этих групп, делается вывод, что, очевидно, существуют долговременные социальные связи между членами группы. Это открытие противоречит принятому мнению, что городские бродячие собаки асоциальны и не формируют стабильных социальных групп. Методологические проблемы могли являться причиной недостоверности более ранних утверждений о том, что городские бродячие собаки не являются социальными. Это произошло по причине неверного использования модели Пуассона для оценки степени социальной организации. Предполагается, что безнадзорные-бездомные собаки, подобно большей части собачьих, для которых существует достоверная база данных, обладают поведенческой пластичностью, позволяющей им приспосабливать свою социальную структуру к требованиям преобладающих экологических ограничений.
В последние годы, популярные и научные тексты демонстрируют растущую обеспокоенность проблемами, вызываемыми большими популяциями неконтролируемых домашних собак в городской и пригородной среде. Эта обеспокоенность вызвала к жизни ряд исследований по естественной истории городских бродячих собак в городах США (Beck, 1971, 1973; Fox et al., 1975; Bekoff , 1979; Westbrook and Allen , 1979; Rubin and Beck , 1982; Berman and Dunbar , 1983; Daniels , 1983 a , b ; Lehner et al ., 1983; Reid et al ., 1984) и в других районах мира ( Oppenheimer and Oppenheimer , 1985; Fox , 1978; Ghosh et al ., 1984; Carr , 1985). Вариации и непоследовательность в сборе и анализе информации делают сравнение этих исследований сложным или невозможным. Также были выявлены результаты, противоречащие друг другу. Особенно, картина, представляющая городских безнадзорных собак асоциальными животными, неспособными к установлению постоянных социальных связей (например, Beck, 1973; Berman and Dunbar, 1983; Daniels, 1983a). Эта идея, упорно воспроизводимая в литературе на протяжении более 10 лет, находится в конфликте с другими исследованиями, которые показывают, что городские собаки формируют стабильные социальные группы (Fox et al., 1975; Carr , 1985).
С августа 1981 по январь 1982 года я провел исследование по это-экологии городских безадзорных собак в Валенсии, городе с 800 000 населения на средиземноморском берегу Испании. Выбранные популяционные параметры были измерены и проанализированы в контексте других схожих работ. В дополнение, были проведены наблюдения поведения в некоторых группах собак для исследования их социальной динамики (Font, 1982). Эта работа представляет данные исследований плотности популяции безнадзорных собак Валенсии и соотношению полов в ней, а также результаты анализа паттернов активности (поведения), размеров участков обитания и территории, выявленных по наблюдениям одной группы, состоявшей из 4 бездомных собак. В заключение, рассмотрен вопрос, являются ли бездомные собаки действительно социальными животными.
МЕТОДЫ
Размеры и структура популяции.
Данные по количеству безнадзорных (т.е. неходящихся вне контроля человека) собак, их пространственной локализации, половом соотношении и приблизительном возрасте были собраны во время периодических учетов по 7 фиксированным учетным маршрутам, расположенных на всей территории города. Для прохождения маршрутов (в течение последовательных 9 дней) использовался маленький мотоцикл, двигавшийся со скоростью около 15 км в час. Маршруты-трансекты были расположены в разных типах городской среды от бедных, экономически депрессивных кварталов до жилых районов среднего класса. Плотность была рассчитано по фотографическому методу мечения-повторного отлова описанному Беком ( Beck , 1973). В этом методе фотография заменяет реальный повторный отлов животного.
Фокус-группа.
Предварительные наблюдения показали, что группы бездомных собак иногда могут оставаться стабильными на протяжении длительных периодов времени. Эти группы демонстрируют иерархическую организацию и совместную защиту территории, что типично для многих диких псовых. Поведение нескольких таких групп было анализировано для того, чтобы проследить их социальную динамику. Одна из таких групп, состоящая из 2 взрослых самцов, 1 молодого самца и 1 самки, была изучена подробно. Все собаки в группе были « feral » (т.е. бездомными) беспородными дворнягами. Их вес по оценке – от 15 до 40 кг. Все были в удовлетворительном физическом состоянии и анализ экскрементов не выявил наличия внутренних паразитов. В начале исследования самка была беременной и родила 3-х щенков, которые были разобраны местными жителями.
На изучение этой группы было затрачено 150 часов наблюдений. Наблюдения были распределены на 3-часовые периоды - так, чтобы покрыть весь суточный цикл активности. Во время каждого периода наблюдений сочетание наблюдений как над собаками по выбору, так и над собаками из фокус-группы (Altmann, 1974a) предоставляло данные по суточной активности собак, размерам участка обитания и территории, типам укрытий, источникам корма и воды и социальному поведению. Паттерны суточной активности были реконструированы посредством усреднения времени, которое каждая особь затрачивала на сон, нахождение в положения сидя или лежа, на протяжении всего периода, в который особь входила в фокус группу. Участок обитания и маршруты передвижения были изучены посредством нанесения на карту местонахождения животных с 10-минутными интервалами. Размер участка обитания был оценен посредством построения на карте выпуклого многоугольника, соединяющего самые удаленные точки встречи животного, и вычисления его площади, исключая внутренние строения и огражденные забором территории, на которые собаки не имели доступа. Были также измерены территория и центральная зона (область ядра).
РЕЗУЛЬТАТЫ
Данные о популяции.
Соотношение полов 2 самца на 1 самку наблюдалось во всех районах изучения ( N = 94 самца, 44 самки) и находится в соответствии с предыдущими исследованиями городских бездомных собак (Beck, 1973; Westbrook and Allen, 1979; Daniels, 1983a). Так как большинство бездомных собак были когда-то домашними, ( это характерно для населенных пунктов с ситуацией, над которой в целом не потерян контроль и самовоспроизводство играет ограниченную роль в поддержании численности бездомных собак – В.Р. ) Дэниелс (Daniels, 1983a) предположил, что такое соотношение отражает выбор в пользу самцов при обзаведении собакой. Однако, отношения полов с сильным преобладанием самцов также были обнаружены среди других видов собачьих (Kleiman and Brady, 1978).
Плотности популяции в 7 исследованных районах варьируют от 127 до 1304 бездомных собак на км². Более высокие плотности имеют тенденцию наблюдаться в наиболее экономически депрессивных районах, особенно тех, где обильны пустыри и постоянные мусорные свалки. Примерно одна треть земли в обычной городской зоне занята зданиями и огражденными заборами площадками, которые не доступны для использования безнадзорными собаками. Они были исключены из расчетов плотности и размеров участков обитания. Эти особенности городской топографии не были учтены при исследованиях в Балтиморе и Ньюарке, где были показаны плотности 232 и 138-204 (интервал значений, три исследованных участка) собак на км², соответственно (Beck, 1973, 1975; Daniels, 1980, 1983a). ( Это в целом справедливое замечание, однако, попытки при учетах вычислять реально доступную для собак площадь вызовет усложнение исследований. Кроме того, относительная доступность пространственных ресурсов является одной из характеристик типа городской среды, поэтому, на наш взгляд, достаточно указывать, к какому типу или категории застройки относится полученное значение плотности. В противном случае в очень плотной застройке, где реально обитает всего несколько собак на кв. км, можно получить сверхвысокие показатели плотности, вводящие в заблуждение сторонних читателей. Сравнение с другими типами среды не будет столь наглядным – В.Р.)
Фокус-группа: участок обитания, территория и паттерны активности.
Таблица 1 показывает размеры участка обитания для группы из 4-х бездомных собак, также как и для другой группы, состоящей из 2 взрослых самцов, которые также были изучены в Валенсии. Представлены для сравнения литературные данные о размерах участка для некоторых групп разного размера и одиночных собак. В некоторых случаях размеры участка вычислялись как полная площадь многоугольника, чьи углы представляют самые отдаленные места встреч. В других – присутствие зданий и огороженных территорий принималось во внимание, и они исключались из расчета («коридорный или линейный участок обитания»; Berman and Dunbar, 1983). Участки обитания групп бездомных собак в сельской местности на один или два порядка больше, чем приведенные здесь ( Scott and Causey , 1973; Nesbitt , 1975; Carr , 1985), возможно за счет более широкого распределения пищевых ресурсов.
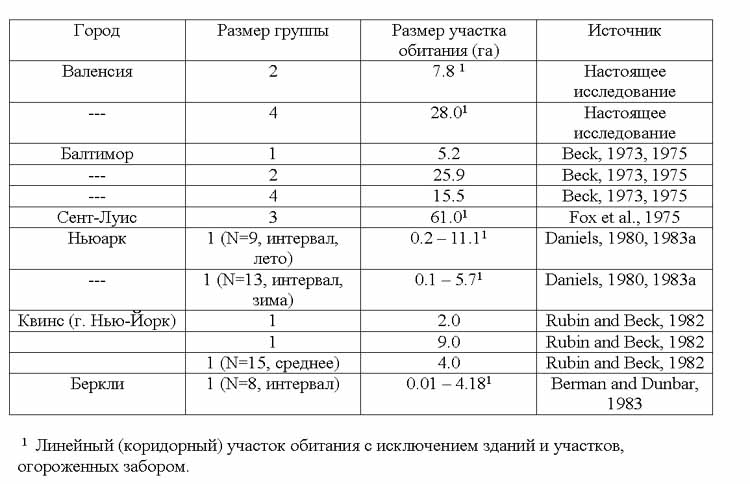
Таблица 1. Участки обитания двух групп бездомных собак, изученных в Валенсии, и оценки размеров участков
обитания других групп (из литературных источников).
В нашем исследовании было выявлено, что использование пространства было не совсем одинаково для разных членов группы, хотя существовало значительное соответствие-перекрывание. Таким образом, групповой участок обитания был вычислен методом суперпозиции (накладывания на карте) участков отдельных членов группы и вычислением размеров площади многоугольника, полученного соединением наиболее отдаленных мест встречи каждого животного.
Размеры «территории» или той части участка обитания, которая активно защищается от чужаков ( Jewell , 1966), были получены для группы из 4 собак путем фиксирования точек, где имели место агрессивные стычки членов группы с потенциальными «нарушителями». . Границы территории было принято считать совпадающими для всех членов группы, так как они все участвовали в этих актах территориальной защиты. Во всех зафиксированных стычках, «нарушители», посягающие на этот эксклюзивный участок, были отогнаны. Однако, владельческие собаки местных жителей, с которыми эти 4 собаки были индивидуально знакомы, не воспринимались как чужаки. Некоторые из этих местных жителей были иногда замечены за передачей пищевых подачек собакам. Групповая территория состояла из пустыря площадью 2 гектара, на котором были заросли кустов и свалки. Меньшая ядерная зона (ядро территории) 0.65 га – область, где произошло 60% от всех встреч наблюдаемых собак, в ее пределах находились излюбленные индивидуальные места отдыха. Несмотря на то, что не было собрано никаких количественных данных по запаховому мечению, очевидно, что наиболее частое размещение точек мечения мочой и эксрементами было вдоль границ территории.
Активность 4 животных в группе была наибольшей ранним утром (7.00 – 10.00) и ранним вечером (17.00 – 21.00). Третий пик активности соответствовал длительным одиночным перемещениям и был отмечен после полуночи. Изучение активности в других исследованиях было основано на подсчете поголовья собак, наблюдавшихся в разные интервалы времени в течение суток. Несмотря на различные методы, использованные для исследования паттернов активности, большинство исследований отмечает бимодальное распределение, с пиками активности утром и в конце дня или начале вечера. В течение типичного дня собаки проводят 18 часов отдыхая (т.е. спят, лежат или сидят) внутри территориальных границ. Когда утром наступает первая фаза активности, собаки были часто замечены в демонстрации «утренних приветствий» - церемонии, состоящей из длительных последовательностей взаимного социального изучения (Fox et al ., 1975).
Агонистические взаимоотношения среди членов группы были очень редки, и ритуализованные позы доминирования или подчинения наблюдались только в нескольких случаях. Это дает основания для утверждения, что основа коммуникации среди членов группы имеет весьма изощренную природу и основана на взаимном распознавании (Fox et al., 1975). Иерархия доминирования была реконструирована на базе этих немногих агрессивных инцидентов. Крупнейший самец был доминантом по отношению к другим 2 кобелям. Из них самый молодой занимал низшую ступень в социальной иерархии, будучи подчиненным по отношению ко всем остальным членам группы. Никаких агрессивных взаимоотношений не наблюдалось между самкой и взрослыми самцами, хотя они были замечены в отступлении, когда сука претендовала на пищу или места лежки.
ОБСУЖДЕНИЕ
Социальны ли городские собаки?
Результаты этого исследования соответствуют во многих отношениях предыдущим исследованиям популяций городских безнадзорных собак. Существует одно расхождение в отношении вопроса: формируют ли собаки стабильные социальные группы. Результаты нашего исследования противоречат широко принятым представлениям, что городские безнадзорные собаки – «асоциальные» животные (например, Scott and Fuller, 1965; Beck, 1973; Kleiman and Brandy, 1978; Berman and Dunbar , 1983; Daniels , 1983 a ) и не противоречат другим исследованиям, показавшим, что существуют длительные связи внутри групп городских собак. (например, Fox et al., 1975; Carr , 1985). Члены социальных групп, исследованных в Валенсии, всегда наблюдались на участке обитания передвигающимися поодиночке или, иногда, парами. ( Видимо, «степень сплоченности» данных, возможно сборных, групп невелика. – В.Р. ). Однако, они защищали общую территорию, имели иерархию доминирования, делились пищей, и членство в группе оставалось неизменным еще долго после окончания исследования (неопубликованные наблюдения). Это дает значительные основания полагать, что в группе существуют долгосрочные связи. К сожалению, популяция, где особи ищут пищу поодиночке, внешне напоминает популяцию, где социальные группы отсутствуют. Вейзер и Джонс (Waser and Jones, 1983) указывают, что особь, занимающаяся фуражировкой в одиночестве, не обязательно уменьшает количество учтенных особей и может жить как член сложной социальной сети. Альтернативное объяснение – то, что стабильные социальные группы – это особенность одних популяций, но не других. ( Вывод в общем случае верный – В.Р.) Однако, перепроверка методов, использованных в ранних работах может привести к выводам, что утверждения о асоциальности обязаны своим происхождением методологической ошибке.
Исследования, находящиеся под вопросом, включают в себя регистрацию частоты встречи групп разных размеров во время учета и сравнение этого распределения с теоретическим распределением частот ( Beck , 1973). Процедура требует выбора определенного критерия для идентификации «группирования» в популяции. Этот критерий может быть основан на пространственной близости животных (например, животные в пределах 10 метров друг от друга приписываются к одной группе, Berman and Dunbar , 1983). Или на комбинации временных и пространственных параметров (например, животные, «остающиеся вместе на протяжении, по крайней мере, 1 мин.» составляют группу; Daniels , 1983 a , p . 343). Затем проводится учет в популяции с фиксированием частоты встречи групп, выявленных по данным критериям. Получающееся распределение частот встреч групп разных размеров сравнивается (с использованием критерия «хи – квадрат», χ 2 ) с ожидаемым распределением частот, сгенерированных с помощью пуассоновского процесса с усечением значения ноль. Исключение нуля необходимо, так как невозможно существование группы с нулем животных в ней. Основанием этого способа служит допущение, что встречаемость групп разных размеров при случайном распределении животных в пространстве соответствует усеченному в нуле распределению Пуассона. Так как животные в этом случае, скорее всего не будут группироваться чаще или реже, чем по простой случайности, наблюдаемое распределение размеров групп, соответствующее модели Пуассона, будет принято как свидетельство того, что эти группы безнадзорных собак - результат случайных, произвольных встреч животных, свободно перемещающихся по своим участкам обитания, то есть не представляют «истинных» социальных групп.
Этот метод был использован по отношению к другим видам (например, кенгуру; Caughley , 1964) и обнаружил определенные недостатки. Прежде всего, должно быть ясно, что использование критерия пространственной близости приводит к получению данных, неподходящих для выводов, если члены группы – одиночные фуражиры. Также следует заметить, что случайное формирование групп может иметь мало общего с социальностью. Нет сомнений, что согласно некоторому произвольно выбранному критерию, группы людей, которые формируются на вечеринке с коктейлем, будут близко соответствовать распределению Пуассона или некоторым другим теоретическим моделям (Cohen, 1971). Мы даже сможем считать обоснованным причисление этих групп к случайным. Однако, заключение из этого, что люди – асоциальные животные, будет явной ошибкой. Кроме того, использование модели для установления социальности у безнадзорных собак некорректно по следующим причинам. Процедура, описанная выше, сравнивает ожидаемые и наблюдаемые распределения размеров групп. Она не берет в расчет пространственное распределение животных. Могут возразить, что пространственное распределение здесь и не рассматривается, рассматривается только распределение размеров групп. Однако, так как группы определяются по критерию пространственной близости особей, сравнение с моделью Пуассона просто некорректно. Вне зависимости от действительного распределения животных в пространстве, всегда можно подобрать такой критерий дистанции, что распределение групп, выделенных согласно ему, соответствовало бы распределению Пуассона. Например, если выбрана слишком маленькая критическая дистанция, можно ожидать, что размер большинства групп будет равен единице. Мы тогда бы пришли к выводу, что наши животные в высшей степени рассеяны, регулярным образом распределяясь посредством какого-то механизма территориального поведения. Наоборот, если выбрана слишком большая дистанция, то обнаружится: большинство животных будет входить в крупные группы. Между этими крайними ситуациями всегда может быть выбрана та дистанция, которая даст результат, соответствующий распределению Пуассона. Если в качестве критерия будет избрана эта дистанция, то мы придем к выводу, что животные формируют группы случайным образом. Таким образом, насколько близко наше распределение размеров групп соответствует пуассоновскому, будет зависеть от двух факторов: 1. нашего первоначального выбора дистанции, 2. распределения животных в пространстве. Это аналогично проблеме, знакомой экологам - выбора «правильного» масштаба для определения паттернов распределения растений и животных (т.е. групповое, случайное или равномерное). Для исследователя, пытающегося анализировать социальную систему безнадзорных собак, неизвестны два параметра: 1. «истинная» наибольшая дистанция, в пределах которой двух животных можно считать социальными, 2. истинное распределение животных в пространстве. Несущественно, какую дистанцию мы выберем, так как не зная закономерности распределения животных в пространстве (например, расстояния между животными в группе или расстояния между группами) мы не имеем оснований сравнивать с моделью пуассоновского распределения. ( То есть нужно учитывать реальные эко-этологические параметры структуры популяции, а не использовать только произвольные формальные критерии – В.Р. ). Помимо этого, хотя определенные группы могут при определенных обстоятельствах адекватно описаны распределением Пуассона (Cohen, 1975), не было бы верным c делать вывод, что все группы в наблюдаемом распределении – случайны.
Приняв во внимание эти аргументы, заявления о несоциальности бездомных собак можно ставить под вопрос. Социальные группы могли существовать в исследованных ( американцами –В.Р. ) популяциях, но не были замечены из-за примененной методики и тенденции приравнивать способность собираться в скопления к социальности. Опираясь на свой опыт, я утверждаю, что никакие статистические манипуляции не могут заменить тщательных и продолжительных наблюдений над индивидуальными животными и группами животных при исследованиях социальной структуры.
Даже если члены социальных групп - как те, о которых упомянуто в этой работе, передвигаются независимо по своим участкам обитания, бродячие собаки в сельской местности демонстрируют определенную тенденцию к формированию стай (Scott and Causey, 1973; Nesbitt, 1975; Carr, 1985). Можно задать вопрос, является ли одиночная фуражировка адаптацией к жизни в городе или продуктом «неправильного» социального поведения?
Волки, делящие с собаками одинаковый набор генов, по наблюдениям Мича (Mech, 1970), живут стаями до 36 особей. Они способны приспосабливаться к изменениям в обилии и распределении своих пищевых ресурсов путем перехода от одиночного к стайному существованию (Fox, 1974). То же самое справедливо и в отношении большинства диких псовых, для которых получены адекватные данные (Kleiman and Brady, 1978). Городские безнадзорные собаки питаются главным образом за счет отбросов и иногда от подачек местных жителей (Beck, 1973; Font, 1982; Daniels, 1983a). Отбросы -мозаично распределенный в пространстве пищевой ресурс. Они доступны собакам в виде маленьких «кусочков», широко разбросанных по их участкам обитания. Такое распределение пищевых ресурсов склоняет к фуражировке на индивидуальной основе или маленькими группами (Altmann, 1974b; Fox et al., 1975). Высоко интегрированные стаи волков и других псовых видимо возникли как адаптация к охоте на крупных травоядных (Kleima n and Eisenberg , 1973; Fox , 1975). Однако, городские собаки не зависят больше от совместной охоты для пропитания. Более того, формирование стай в городской среде может даже быть контрадаптивно, так как большие группы собак редко терпимы людьми и являются объектом «охоты» со стороны муниципальных ловцов. ( Если, конечно, в городе есть контроль содержания и эффективная служба отлова. Как показывает опыт России, большие стаи в городах возможны в местах с укрытиями (за заборами), обильной пищевой базой и слабым (по сравнению с Америкой) муниципальным контролем. Размер стаи в этом случае действительно не играет никакой адаптивной роли в деле поиска пищи и зависит почти только от сочетания благоприятных условий, позволяющих реализовываться наследственным предпосылкам социального поведения – В.Р. ). В сельской местности, с другой стороны, кооперация всех членов группы может быть востребована для умерщвления большой добычи. Наблюдения над собаками-париями в Индии обнаружили новые особенности поведенческой пластичности. Здесь собаки с неперекрывающимися территориями иногда собираются вместе для охоты на оленя или для отпугивания чужаков с общих фуражировочных/охотничьих участков (Fox, 1975).
ВЫВОДЫ
Из нашего исследования следуют четыре основных заключения.
1. Для того, чтобы можно было сравнивать исследования, нужно достичь согласия относительно методик, используемых для сбора и анализа информации по безнадзорным собакам (например, расчета плотности и размеров участков обитания, паттернов суточной активности).
2. Сравнивание встречаемости групп разных размеров с усеченным в значении ноль распределением Пуассона как способ оценки социальных тенденций в поведении собак – не является методом, подходящим для данной задачи. Следовательно, утверждения, что городские безнадзорные собаки асоциальны – видимо, не имеют оснований.
3. Результаты этого исследования показывают, что городские бездомные собаки живут в сложной интерактивной сетевой структуре со всеми атрибутами постоянных социальных групп.
4. Разница между группами городских и загородных собак объяснима на основе распределения пищевых ресурсов. ( Но шире - не полностью только за счет распределения пищи – В.Р.) Пластичность, демонстрируемая социальной системой собак имеет параллели с тем, что наблюдалось у диких псовых.
ИСТОЧНИКИ
Altmann, J., 1974a. Observational study of behavior: Sampling methods. Behaviour, 49:227-267.
Altmann, S. A., 1974b. Baboons, space, time, and energy. Am. Zool., 14:221-248.
Beck, A. M., 1971. The life and times of Shag, a feral dog in Baltimore . Nat. Hist., 80:58-65.
Beck, A. M., 1973. The Ecology of Stray Dogs: A Study of Free-ranging Urban Animals. York Press, Baltimore , MD , 98 pp.
Beck, A. M., 1975. The ecology of "feral" and free-roving dogs in Baltimore . In: M. W. Fox (Editor) , The Wild Canids: Their Systematics, Behavioral Ecology and Evolution. Van Nostrand Reinhold , New York , pp. 380-390.
Bekoff, M., 1979. Scent-marking by free-ranging domestic dogs. Biol. Behav., 4:123-139.
Berman, M. and Dunbar , I. , 1983. The social behaviour of free-ranging suburban dogs. Appl. Anim. Ethol., 10: 5-17.
Carr, G. M., 1985. Behavioural ecology of feral domestic dogs ( Canis familiaris) in Central Italy . XIX Int. Ethological Conference, Toulouse , France , abstract 267.
Caughley, G., 1964. Social organization and daily activity of the red kangaroo and the grey kangaroo. J. Mammal., 45:429-436.
Cohen, J. E., 1971. Casual Groups of Monkeys and Men. Harvard University Press, Cambridge , 175 pp.
Cohen, J. E., 1975. The size and demographic composition of social groups of wild orang-utans. Anim. Behav., 23:543-550. Daniels, T. J., 1980. The social behavior of free-ranging urban dogs. Master's Thesis, Ohio State University.
Daniels, T. J., 1983a. The social organization of free-ranging urban dogs. I. Non-estrous social behavior. Appl. Anim. Ethol., 10: 341-363. Daniels, T. J., 1983b. The social organization of free-ranging urban dogs. II. Estrous groups and the mating system. Appl. Anim. Ethol., 10: 365-373. Font, E., 1982. Eto-ecologia del perro vagabundo. Master's Thesis, University of Valencia ( Spain ). Fox, M. W., 1975. Evolution of social behavior in canids. In: M. W. Fox (Editor), The Wild Canids: Their Systematics, Behavioral Ecology and Evolution. Van Nostrand Reinhold, New York , pp. 429-460.
Fox, M. W., 1978. The Dog: Its Domestication and Behavior. Garland STPM Press, New York , 296 pp.
Fox, M. W., Beck, A. M. and Blackman, E., 1975. Behavior and ecology of a small group of urban dogs (Canis familiaris). Appl. Anim. Ethol., 1:119-137. Ghosh, В ., Choudhuri, D. K. and Pal, В ., 1984. Some aspects of the sexual behaviour of stray dogs, Canis familiaris. Appl. Anim. Behav. Sci., 13:113-127. Jewell, P. A., 1966. The concept of home range in mammals. Symp. Zool. Soc., London , 18:85-109. Kleiman, D. G. and Brady, C. A., 1978. Coyote behavior in the context of recent canid research: Problems and Perspectives. In: M. Bekoff (Editor), Coyotes: Biology, Behavior and Management. Academic Press, New York , pp. 163-188.
Kleiman, D. G. and Eisenberg, J. F., 1973. Comparisons of canid and felid social systems from an evolutionary perspective. Anim. Behav., 21:637-659.
Lehner, P. N., McCluggage, C, Mitchell, D. R. and Neil, D. H., 1983. Selected parameters of the Fort Collins , Colorado , dog population, 1979-80. Appl. Anim. Ethol., 10:19-25.
Mech, L. D., 1970. The Wolf: The Ecology and Behavior of an Endangered Species. Natural History Press, Garden City, New York , 384 pp.
Nesbitt, W. H., 1975. Ecology of a feral dog pack on a wildlife refuge. In: M. W. Fox (Editor), The Wild Canids: Their Systematics, Ecology and Evolution. Van Nostrand Reinhold , New York , pp. 391-395.
Oppenheimer, E. C. and Oppenheimer, J. R., 1975. Certain behavioral features in the pariah dog (Canis familiaris) in West Bengal . Appl. Anim. Ethol., 2: 81-92. Reid, J. В ., Chantrey, D. F. and Davie , C, 1984. Eliminatory behaviour of domestic dogs in an urban environment. Appl. Ani m. Behav. Sci., 12:279-287. Rubin, H. D. and Beck, A. M., 1982. Ecological behavior of free-ranging urban pet dogs. Appl.
Anim. Ethol, 8:161-168. Scott, J. P. and Fuller, J. L., 1965. Genetics and the Social Behavior of the Dog. University of Chicago Press, Chicago, 468 pp. Scott, M. D. and Causey, K., 1973. Ecology of feral dogs in Alabama . J. Wildl. Manage., 37:253-265. Waser, P. M. and Jones, W. Т ., 1983. Natal philopatry among solitary mammals. Q. Rev. Biol., 58: 355-390.
Westbrook, W. H. and Allen, R. D., 1979. Animal field research. In: R. D. Allen and W. H. West-brook (Editors), The Handbook of Animal Welfare. Garland STPM Press, New York , pp. 148-167.
Перевод и подготовка текста В.А. Рыбалко